| Issue |
BIO Web Conf.
Volume 24, 2020
International Conferences “Plant Diversity: Status, Trends, Conservation Concept” 2020
|
|
|---|---|---|
| Article Number | 00067 | |
| Number of page(s) | 5 | |
| DOI | https://doi.org/10.1051/bioconf/20202400067 | |
| Published online | 21 September 2020 | |
The genus Adoxa L. (Adoxaceae Trautv.) in the flora of Russia
Transbaikal State University, 672039 Chita, Russian Federation
* Corresponding author: This email address is being protected from spambots. You need JavaScript enabled to view it.
Abstract
In this paper, species of the genus Adoxa L. were analyzed that occur in the flora of Russia. We noted that three species have been found on the territory of Russia: Adoxa moschatellina L., Adoxa orientalis Nepomn. and Adoxa insularis Nepomn. A. moschatellina has a circumpolar areal. The species A. orientalis rarely occurs in Far East. It was determined that this species has more primitive flower structure as compared with A. moschatellina. The lateral flowers of A. orientalis in the inflorescence are trimerous (Ca(3)CO(6)A6G(3)),, whereas anther filaments are integral. On islands Kunashir and Sakhalin, we noted the presence of A. insularis whose flower structure has signs of more complete organization. In A. insularis, petals are oval, with a pointed tip, and have straight veinlets; A. insularis possesses 3- to 4-fold fewer nectar glandules, which are located directly on the petals’ surface, rather than in depressions on the petals as in A. moschatellina. Anther filaments in A. insularis are longer, and the angle between cleft filaments in A. insularis is 30–40° (rather than 90° or more as in A. moschatellina). It was concluded that in Adoxa species, there has been an obvious process of evolutionary reduction of flower structure.
© The Authors, published by EDP Sciences, 2020
 This is an Open Access article distributed under the terms of the Creative Commons Attribution License 4.0, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License 4.0, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
For a long time, the Adoxaceae family has been regarded as monotypic and represented by one species, Adoxa moschatellina L. [1]. This species has a circumpolar areal and is widespread in moderate and cold regions of the Northern hemisphere and in mountain forests of its tropical zone. In Russia, this species inhabits conifer and mixed forests; alder, aspen, and willow thickets; fontinal banks; and sites near cliff piedmonts; in mountains, it reaches the Alpine belt. In the North, it occurs also on the islands of the Arctic Ocean [1].
A. moschatellina is a short-rooted semirosular herbaceous plant whose flowers are gathered into a capitate inflorescence. In this species, the ratio of segments of the apical flower is usually divisible by two and expressed by the formula Ca(2)CO(4)A4G(4). Lateral flowers are mostly pentamerous. Their structure is expressed by the formula Ca(3)CO(5)A5G(5) (hereafter, for brevity, flower formulas will be presented as 3.5.5.5.). When studying the variation of A. moschatellina flowers, we found [2] that most of its lateral flowers (73) 85-92% have typical structure, with a flower segment ratio of 3.5.5.5. Besides, some inflorescences contain flowers some segments of which are even more reduced and conform to ratios 3.5.5.4, 3.4.4.5, 3.4.5.4, 3.4.4.4, 2.4.4.4, and 2.4.3.4. Additionally, flowers with segments featuring divisibility by three (3.6.6.6, 3.6.6.5, 3.6.6.4, and 3.6.5.4) can occur. This variation indicates the gradual nature of the processes of evolutionary reduction. Anther filaments in the flowers of this species are completely cleft (Fig. 1).
On the territory of Russia, two new species of Adoxa have been found, which have a limited habitat area [3]. A. orientalis Nepomn. is a novel species that was described by us [4] using herbarial material collected in 1912 in Amur Oblast, near Zeya-Pier, meteorological station Pikan, on a stream bank, on hillocks, A.P. Archangelskay. In Chita Oblast, this species was collected in 1909 at two additional locations (Nerchinsko-Zavodsky District, basin of Argun River, Urumkana River system, right bank of Orocha River, upward from Kamenucha creek valley, in communities of mhas along the ravine bed, I.I. Krasheninnikov; and Nerchinsko-Zavodsky District, watershed between Shilka River and CherniyUryum River, bottom of Morgorundra creek valley, N. B. Blagoveshchenskay, G.I. Pavlovskay). In the herbarium of the Federal Scientific Center of the East Asia Terrestrial Biodiversity(Far Eastern Branch of Russian Academy of Sciences), there is a herbarial specimen collected in Khabarovsk Krai in the basin of Uda River, in a small-leaved forest on a bank of Ugokhan River, U.A. Doronina. In 2013-2014, the species was collected in Primorsky Krai in Ussurisky Nature Reserve, in the upper parts of Artemovka River and BolshayaSuvorovka River [5].
In 2002, we for the first time [6] found a small population of this species in the Gazimuro-Zavodsky District of Transbaikal Krai, in the basin of Budyumkan River, near stream Typkuraucha, in Tagaiskaya creek valley, 3 VI 2001. A. orientalis was collected near a natural spring at the bottom of a deep narrow creek valley that was bounded by high steep ridges. The main type of vegetation there is moss-covered larch forests (Laricetum sphagnatum). Here, according to descriptions by I.M. Krasheninnikov (1954) [7], “peculiar communities of mhas’ can be found where Ranunculus lapponicus L. occurs here and there, and where a thick moss cover (sometimes pure sphagnum, sometimes with a touch of other mosses: e.g., Hylocomium proliferum, Rhytidium rugosum, Camptothecium nitens, Ptilidium ciliare, Aulacomnium palustre and Thuidium recognitum) reaches tens of centimeters in thickness”.
In lower reaches of the Typkuraucha stream (where the creek valley starts to widen), due to substantial cross-section width, lowering of the surrounding elevations, and the resultant increased aeration and illumination, more favorable conditions are formed for the development of other types of vegetation. In this region, the area of larch forests is decreasing. On southern slopes of creek valleys, steppe associations of plants develop, and sometimes, Quercus mongolica Fisch. ex Ledeb. occurs. On the northern slopes, birch forests are common. Valley bottoms are mostly occupied by meadows on overhumidified soils. Our study showed that in such plant associations, along riverbeds, only A. moschatellina grows at present, whereas A. orientalis does not occur in such plant associations. Thus, the modern confinedness of A. orientalis growth to deep “creek valleys” is an obvious consequence of higher humidity, strong inumbration, and the nearby location of permafrost, which substantially lowers the soil temperature. In such plant associations, stable coenotic connections are formed, which are also joined by A. orientalis.
In contrast to A. moschatellina, A. orientalis has ternate (rather than double ternate) leaves of the rosette near the root, and their leaflets are located on broad petiolules, which are 5- to 6-fold shorter than those in A. moschatellina. The petiole of the leafstalk is 1.5-2.5 (rather than 0.0-0.7) mm wide, with well-pronounced green wings decurrent along the whole petiole [4]. A. orientalis possesses the most primitive structure of flowers in comparison with all the known species of Adoxa. It is this species that features inflorescence lateral flowers that have a structure corresponding to the formula Ca(3)CO(6)A6G(3), and its anther filaments are integral, and even anthers are sometimes fused (Fig. 2), i.e., retain the traits that are close to the original type [2]. On the other hand, in A. orientalis flowers, gynoecium reduction is more profound (down to three carpels). This finding suggests that in the course of development, the reduction of different segments of the flower is not simultaneous. This combination of traits of different evolutionary significance indicates this species’ antiquity.
The presence of A. insularis Nepomn. is predicated on the conditions of mountainous dark coniferous taiga of Pacific islands (islands Kunashir and Sakhalin) [8]. On Kunashir Island, this species was found 20 km north of the Yuzhno-Kurilsk settlement, natural spring Dobriy, on a hill, in a coniferous forest, in the same location on a river bank in a calla; in the same location on a bank of Filatovka River, in a coniferous forest. 20 VI 1983. O.A. Nepomnyaschaya. On Sakhalin Island, another habitat of this species was noted: Eastern Sakhalin Mountains, Nabilsky ridge, Lopatin Mountain (1000-1200 m high), a belt of cedar carpeting bush, among stones near a melting snow patch 26 VII 1984. O.A. Nepomnyaschaya [2-4].
The described species strongly differs from A. moschatellina (Fig. 1). In A. insularis, the petals (Fig. 3) are oval long narrow, with a pointed tip, and have straight veinlets, as opposed to the broad rounded petals of A. moschatellina, which have a blunt rounded tip and branched veinlets; the corolla tube of A. insularis is half as long as that of A. moschatellina. A. insularis has 3- to 4-fold fewer nectar glandules, which are positioned directly on the surface of petals (Fig. 3) rather than in depressions on the petals as in A. moschatellina (Fig. 1). A. insularis anther filaments are longer, and the angle between cleft filaments in A. insularis is 30-40° (rather than >90°as in A. moschatellina).
It should be pointed out that in A. insularis, the flowering period is very long: lasting from June till the end of July. A. moschatellina growsin all other forest associations of plants on the whole territory of Sakhalin Island, and the flowering period of this species matches the spring season.
Thus, in Adoxa species that grow on the territory of Russia, there is a clear-cut process of evolutionary reduction of flower structure.
References
- B.K. Shishkin, Flora of the USSR. (AN USSR, Moskow-Leningrad, 1958) [Google Scholar]
- O.A. Nepomnyaschaya, J. Bot., 69, 1030 (1984). [Google Scholar]
- O.A. Nepomnyaschaya, Vascular plants of the Soviet Far East. (Nauka, Leningrad, 1988) [Google Scholar]
- O.A. Nepomnyaschaya, J. Bot., 69, 259 (1984) [Google Scholar]
- L.A. Fedina, Komarov readings, LXV (Dalnauka, Vladivostok, 2017) [Google Scholar]
- O.A. Popova, J. Bot., 87, 131 (2002) [Google Scholar]
- I.M. Krasheninnikov, Geographical works. (Geographical Literature, Moscow,1954) [Google Scholar]
- O.A. Nepomnyaschaya, J. Bot., 70, 524 (1985) [Google Scholar]
All Figures
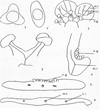 |
Fig. 1 Structure of the Adoxa moschatellina L. flower 1, l’. The calyx of lateral and apical flowers. 2.General view of a petal with stamens. 3.Stamen. 4.Longitudinal sectionof a petal with nectar glandules. 5.Cross-section of a petal with nectar glandules. 6.Cross-section of a petal. n.g.: nectar glandules, m.r.: midrib, l.r.: lateral rib. |
| In the text | |
 |
Fig. 2 Structure of the Adoxa orientalis Nepomn. Flower. 1. Corolla of an apical flower. 2.Corolla of a lateral flower. 3.Gynoecium of an apical flower. 4.Gynoecium of a lateral flower. 5.Stamen. |
| In the text | |
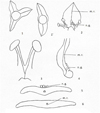 |
Fig. 3 Structure of the Adoxa insularis Nepomn. Flower 1, 1’. The calyx of lateral and apical flowers. 2.General view of a petal with stamens. 3.Stamen. 4.Longitudinal section of a petal with nectar glandules. 5.Cross-section of a petal with nectar glandules. 6.Cross-section of a petal. n.g.: nectar glandules, m.r.: midrib rib. |
| In the text | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.